转自知乎https://www.zhihu.com/question/20264424 科研君的回答
学习和记忆是所有动物适应其生存环境所需要的最基本能力,所以有关学习和记忆的研究一直以来都非常热门。记忆的本质,从不同领域来看有不同的侧重,比如说它的基本生物结构,它的心理学特征和组织还有它的持久性等等,数百年来,虽然有来自生物、心理学领域的科学家甚至哲学家等学者都试图通过不同的研究方法来试图寻找这类问题的终极答案,但是坦白的说,这是个非常大而复杂的问题,而且到目前为止还没有确定的答案,不夸张的说,人类的学习和记忆是现存进化程度最高也是最复杂的生物加工过程。
所以,我在这里只想从我自身的理解和认识上来简化谈谈这个问题(我是医学和生物背景,所以可能不会从计算科学角度去探究这个问题)。因为学习也是记忆,学习的唯一证据就是记忆,所以关于学习的储存其实可以理解成记忆的存储问题,这样一来,再根据题主的问题描述,我将提问总结归纳以下几个方面的问题:1。记忆的本质2.记忆的储存问题3.记忆的提取问题
1.记忆的本质
从神经生物学的角度来看,目前主流观点是“神经元学说”,认为记忆是在神经细胞间联结的动态变化中得到的编码的,换句话来说,记忆可以体现神经细胞之间联结的可变性和可塑性。可以看出来,记忆的形成不是哪个神经元决定的,而是有很多神经元相互连接,相互作用的。记忆按照记忆时程的长短分为瞬时记忆、短时记忆、长时记忆和永久记忆,目前认为各类记忆的脑内机制是不相同的,记忆的分类以及不同记忆类型的比较如图所示。
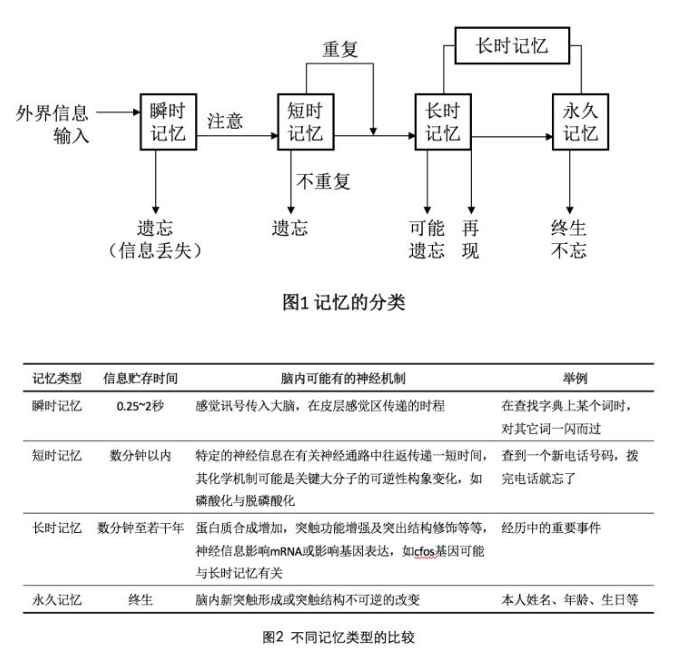
当然,我也一直强调神经科学领域的发展和进展是非常迅猛的,很多观点和结论都在不断被完善和更新。我记得再2016年1月份的一次有关学习与记忆的论坛上,钟毅老师提出过“重复学习不改变记忆的形成,重复改变记忆的遗忘”的观点,也就是说它认为每次学习过程中,第一次产生的是学习,后面的重复都是抑制遗忘。他用果蝇为模型,用行为学、细胞分子学等研究方法阐明自己的观点,这个观点打破了短时程记忆转成长时程记忆大多需要consolidation(巩固)的过程的理论,当然,这也只是一种理论,而且模式动物用的是无脊椎动物果蝇,所以是否再灵长类动物和人类身上也使用也是不得而知的。
深一步的从学习与记忆的分子机制来看,现在的研究认为,记忆源于突触的微小改变,这种变化再全脑都有体现。这样一来,记忆的物质基础的研究范围缩小到突触修饰上,于是科学家先从最简单的无脊椎动物神经系统入手探索记忆的分子机制。这里首先要提到的是20世纪末,美国哥伦比亚大学科学家Eric Kandel成功地建立了海兔地学习和记忆行为模型,这一先驱性实验显示了记忆地存储位点和形成机制,从而系统地解释了学习记忆可能地细胞和分子机制,也因此他获得了2000年诺贝尔奖。这个模型主要研究的是程序性记忆,包括海兔地非联合性学习和海兔地联合型学习,其中非联合型学习又包括“缩鳃反射地习惯化”和“缩鳃反射地敏感化”,而联合型学习也就是指经典地条件反射。这里简单介绍一下它们地细胞机制:
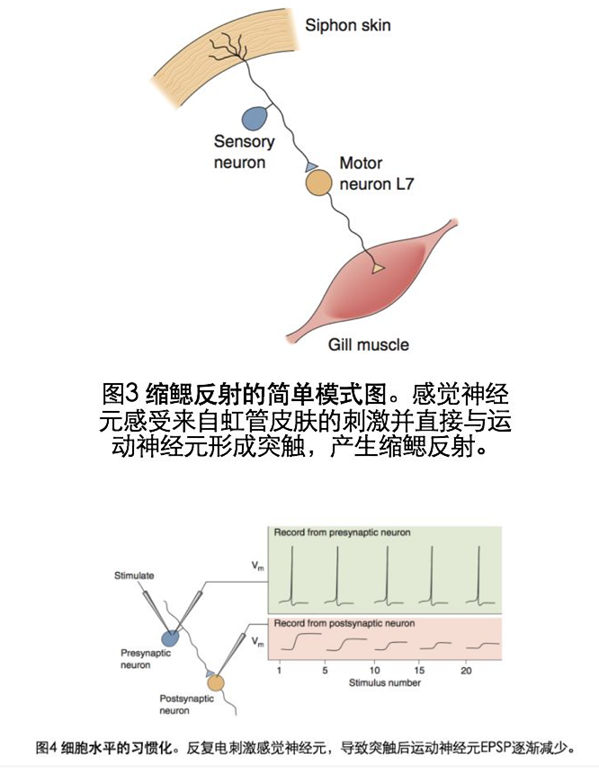
1)海兔地非联合型学习——缩鳃反射地习惯化
如果将水流喷射到海兔地虹管,虹管和鳃会收缩,称为缩鳃反射(图3).Kandal实验的结论是:缩鳃反射的习惯化与突触前修饰有关,反复刺激感觉神经末梢会使神经递质的释放量减少。进一步来讲,钙离子通过电压门控钙离子通道进入神经末梢,但是当钙离子通道反复开放时,他的效能就会降低,钙离子内流减少,神经递质释放量减少。(图4).
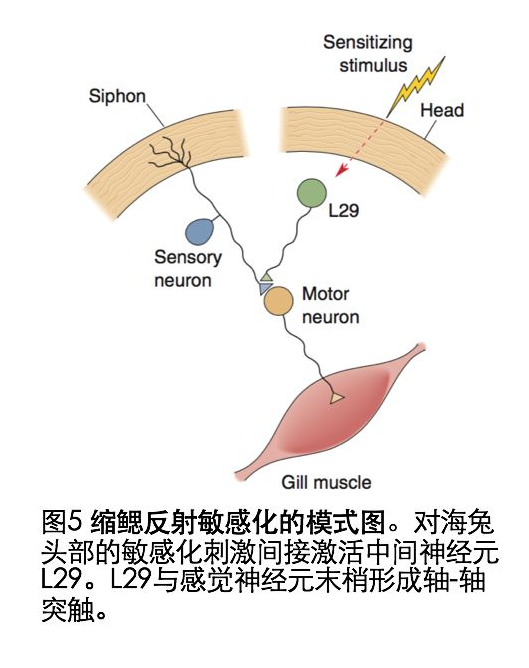
2)海兔的非联合型学习——缩鳃反射的敏感化
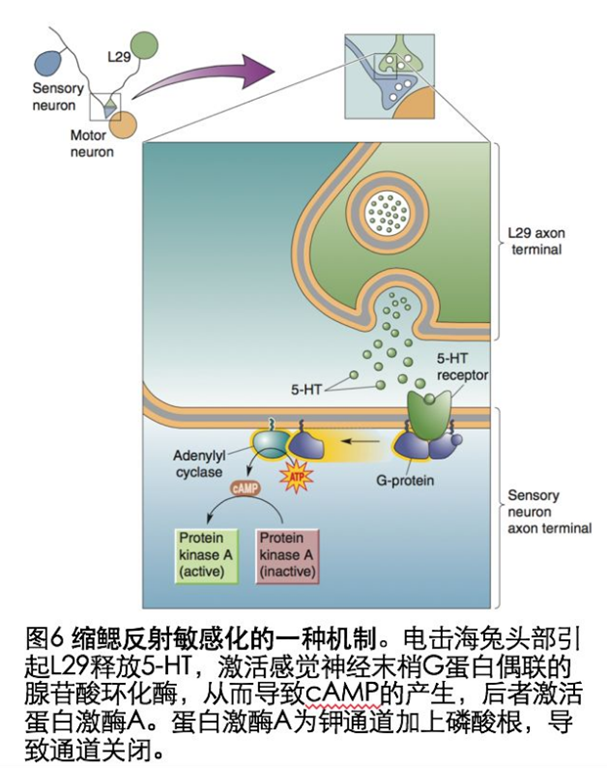
短暂电击海兔的头部,结果导致再刺激虹管引起的强烈的缩鳃反应(图5).由于电击海兔头部会引起L29释放五羟色胺(5—HT),随后激活感觉神经末梢G蛋白耦联的腺苷酸环化酶,随后导致cAMP产生,cAMP可以激活蛋白激酶A(PKA),PKA磷酸化钾离子通道蛋白,导致钾离子通道关闭,突触前动作电位延长,促进动作电位期间钙离子内流增多,神经递质释放量增多(图6).
3)海兔的联合型学习
即非条件刺激是强电击海兔尾部,条件刺激是对虹管的轻微刺激,后来如果将尾部刺激和虹管刺激相结合后,单独刺激虹管后产生的反应要比敏感化引起的反应还要强。从分子水平上的解释是条件刺激表现为钙离子内流,非条件刺激表现为突触前神经末梢腺苷酸环化酶的G蛋白激活,当突触前钙离子冲动与G蛋白耦联受体激活导致cAMP大量生成在时间上恰好重合,或者钙离子冲动稍微较后者快一点时,产生学习,而记忆则在钾离子通道被磷酸化和神经递质释放增加时产生。(图7)
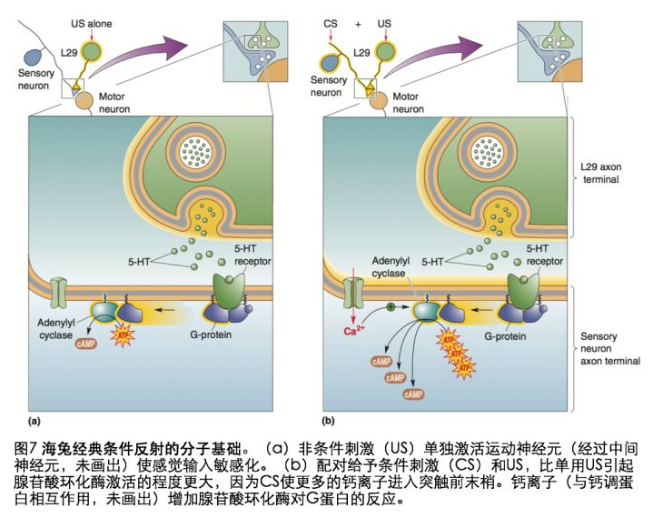
关于用脊椎动物模型来研究哺乳动物的突触可塑性的内容是非常的多,在这里我只提几个哺乳动物中学习和记忆的细胞分子学机制中非常重要的关键词:海马的长时程增强(long-term Potentiation,LTP),突触传递的长时程压抑(long-term depression,LTD),NMDA受体(N-methyl-D-aspartate receptor)。总的来说,药理学和遗传学的研究都表明,海马NMDA受体在突触修饰作用(比如说LTD和LTP)和学习记忆中都有着很关键的作用。
2.记忆的储存
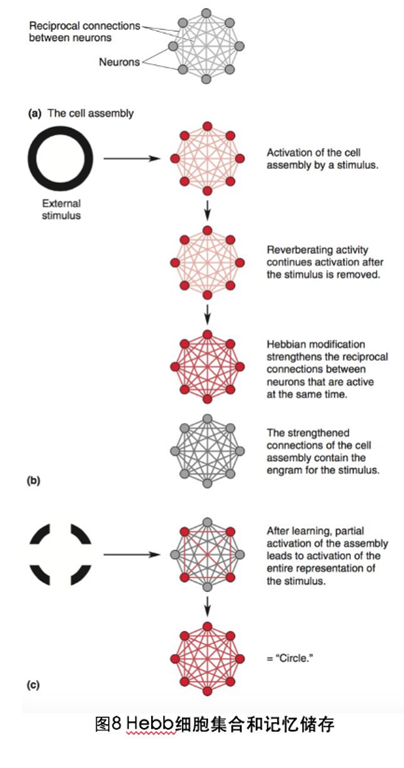
记忆的物质表现或位置可以叫做记忆的痕迹(memory trace),同时也不得不提到加拿大心理学家Donald Hebb,Hebb提出了细胞集合理论:脑内反映外界客观物体,是由被外界刺激激活的所有皮层细胞组成的,这些同时被激活的神经元称作“细胞集合”,假如这些细胞相互连接,细胞集合内的连接持续激活,对外界客观物体的内部反应就能作为短时程记忆始终保存,如果细胞集合能维持激活很长一段时间,那么细胞间相互连接更有效的神经元就会连接在一起,更紧密的连接就会使细胞集合再次兴奋,记忆的巩固就可能发生(图8).简单的讲,他的重要结论包括:
1)仅仅集团内的一部分细胞的破坏并不能消除记忆,记忆的痕迹广泛分布于细胞集合的细胞连接内;
2)记忆痕迹可能包括了有关感觉和知觉的神经元
不管怎么样,目前认为学习和记忆的功能不局限于大脑某一单一的部位,而研究表明,陈述性记忆主要依赖于海马和相关结构,程序性记忆与纹状体有关,而工作记忆的痕迹是广泛分布于大脑内的,另外,长时程记忆被一些人认为是通过新皮层结构的变化来存储的记忆。所以说为了保障我们更好的生存下去,我们的大脑是一直在不停的再重构的。
还有说法是,多数记忆材料都需要经过Joseph LeDoux提到的“短时缓冲装置”,即位于双侧听觉区、视觉区和运动知觉区的暂时存储区域后,然后才能进入工作记忆(一种暂时记忆,比短时记忆要长一些)和之后的长时记忆。而语义记忆、情景记忆、程序记忆、自动记忆和情绪记忆的通路则用于储存和提取长时记忆的信息。其中,情景记忆和语义记忆(两者都属于陈述性记忆)主要储存在海马;程序记忆和自动记忆主要储存在小脑;情绪记忆主要储存在杏仁核。这里再简单解释一下这五种记忆:
1.语义记忆:人们对一般知识和规律的记忆,比如你阅读文献所得到知识的记忆
2.情景记忆:可以理解成关系记忆或者空间记忆,主要指位置信息
3.程序记忆:通常也称为“肌肉记忆”,比如我们学开车、骑自行车等记忆
4自动记忆:也称作为conditioned response memory,表现为特定的刺激对记忆的自动激发,例如说你唱着歌我自动的就接下一句的这种状态
5.情绪记忆:就是关于情绪的记忆。值得一提的是,情绪记忆总是优先于其他任何一种记忆
3.记忆的提取问题
刚才有提到语义记忆、情景记忆、程序记忆、自动记忆和情绪记忆这5条记忆通道,一种说法是,我们很多时候会通过这些记忆通道来激活记忆,开启记忆。具体的举个例子来讲,寒假回家爸爸接我,车里放着CD,当放到《同桌的你》时,我都会不自主地跟着哼(歌曲开启了自动记忆),然后当车经过我初中学习地时候,都会勾起我对美好童年的回忆(通过初中学校这个地点,情景记忆通道被激活),然后我想起每天放学和同学迫不及待开心地蹲在学校门口看商贩贩卖小鸡小鸭和蚕的场景,觉得小时候的我好天真可爱,“噗嗤”一下笑了出来,于是心情大好,突然想下车看看学校门口有什么新变化(情景记忆通路激活情绪记忆,同时由于情绪记忆总是优先于其他记忆的,当情绪记忆主导时,人很容易失去逻辑能力,做出背离理智的决定),我于是来到校门口,发现校门口开了家新的炸鸡店,名字叫“来自星星的炸鸡”,我买了一份尝了后觉得很好吃,突然想起闺蜜说过她也很喜欢吃炸鸡(语义记忆通路打开),于是我微信告诉她校门口新开了一家好吃的炸鸡店,明天约她一起吃炸鸡。
总结
神经科学是一个非常值得探索的领域,我们的大脑大约有1000亿个神经元,但是它们到底是如何工作的,依然是个迷。
目前就我们的了解,记忆能是发生在突触处的,而且各类动物的学习和记忆在细胞和分子机制上都是很相通的。典型的事件是:脑中电活动的改变,然后第二信使分子的变化(如cAMP,钙离子),之后突触蛋白的修饰,这些暂时的变化通过改变突触结构从而成为永久变化,形成长时程记忆。随着神经科学的迅速发展,我相信会有更多更新颖,合理和突破性的理论产生,说不定哪天真发明出让人过目不忘的“记忆面包”呢!
原文地址:https://www.cnblogs.com/yifdu25/p/8454681.html